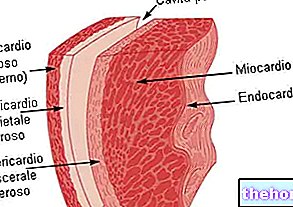
BILJEŠKE O EMBRIOLOGIJI
Embriologija proučava slijed oblika razvoja od zigote do organizma obdarenog svim njegovim organima i sustavima.
U tom smislu vrijedi se sjetiti razlike između razvoja (sukcesije strukturnih i organizacijskih faza sa sve većom složenošću) i rasta, namijenjenog prije svega u kvantitativnom smislu.
U metazoanima kralježnjaka svjedočimo porastu u evolucijskom nizu sve do čovjeka (preko ciklostoma, riba, vodozemaca, gmazova, ptica i sisavaca), pojavi odraslih oblika sve veće složenosti, zbog čega dolazi do komplikacija faza embrionalnog razvoja.
Na početku, zigota, uvijek opremljena rezervnim materijalom, podijeljena je (naknadnom mitozom) na 2, zatim 4, zatim 8 itd. Stanice zvane blastomere, bez rasta, sve dok ne dosegne normalni omjer jezgre / citoplazme vrste .
Ova početna segmentacija može slijediti različite obrasce, ovisno o količini i distribuciji deutoplazme.
U početku je deutoplazma oskudna ("oligolecitna jaja"), pa je segmentacija potpuna i dovodi do vrlo različitih blastomera. Kako se povećava složenost embrija, potrebno je više vremena i materijala prije nego što mu razvoj omogući početak. samostalan život. To zahtijeva povećanje deutoplazme ("telocitna jaja"), koja se nastoji rasporediti u dijelu zigote. To uzrokuje sve veću "anizotropiju", koja je povezana s izmjenama segmentacije, prema dva opća načela:
- Hertwigov zakon kaže da se u mitozi akromatsko vreteno (čiji ekvator određuje ravninu diobe stanica kćeri) nastoji rasporediti u smjeru najveće duljine citoplazme;
- Balfourov zakon kaže da je brzina segmentacije obrnuto proporcionalna količini deutoplazme.
Tada vidimo da je već u ciklostomima i u ribama segmentacija nejednaka, s brzo segmentiranim životinjskim polom (koji će dati gornje strukture embrija) i žumanjčanim stupom koji će sadržavati većinu rezervnog materijala. Ova je tendencija još veća anizotropni kod vodozemaca (u kojima je potrebno pripremiti organe odgovorne za disanje zrakom), u kojima stup žumanjka, iako se polako segmentira, ostaje relativno inertan i završava prekriven stanicama dobivenim iz brzo segmentiranog životinjskog pola. glavni embrionalni stadiji uključuju: zigotu, blastomere, morulu (skup blastomera sličnih kupini), blastulu (morulu s regresiranim unutarnjim stanicama), gastrulu (blastulu u kojoj su stanice s jedne strane invaginirane), u kojoj je primitivna šupljina organizma, s vanjskim staničnim slojem (ektoderm, iz kojeg će prije svega proizvesti živčani sustav) i int erno (entoderm), između kojih će se zatim ubaciti treći sloj (mezoderm). Iz tih slojeva ili "embrionalnih ploča" tada će, u uređenom slijedu, izvesti sva tkiva, organi i sustavi.
Kod još razvijenijih vrsta povećanje deutoplazme (ili "teleta") je takvo da se ne može ni segmentirati. Tako vidimo da segmentacija kod ptica utječe samo na tanki površinski disk, što dovodi do "diskoblastule" i niza pojave koje jamče nastanak embrija na drugačiji način od gore spomenutog.
Daljnje povećanje deutoplazme vjerojatno ne bi bilo učinkovitije, pa se kod sisavaca razvoj i rast do sposobnosti za neovisan život postižu drugim sustavom. Zapravo primjećujemo da se deutoplazma koristi samo u prvim fazama razvoj; tada zametak uspostavlja metaboličke odnose s majčinim organizmom (s pojavom posteljice) i više ne koristi deutoplazmu, čiji se višak eliminira. U tom se trenutku jajašca vraćaju u oligolektit i segmentacija se može vratiti da je potpuna (pa je stoga u ranim fazama slična onoj u "amfioksusa"), ali nakon morule embriogeneza se nastavlja prema najrazvijenijem uzorku ptica, s "blastocistom" nakon koje slijedi implantacija na stijenku maternice, tako da metabolizam embrija osigurava majčinski organizam (putem posteljice), a ne deutoplazma.
EMBRIJSKA DIFERENCIJACIJA
Kad segmentacija zigote omjer jezgre / citoplazme dovede u normalu vrste, rast također mora započeti, paralelno s razvojem. Iz tog razloga počinje metabolizam, pojavom jezgri i sintezom proteina. Tako započeta sinteza proteina posljedica je gena odgovornih za rane faze embrionalnog razvoja. Ti su geni potisnuti tvarima prisutnim u različitim blastomerima životinjskog i telećeg pola. Zauzvrat, proizvodi ovih ranih gena mogu poništiti djelovanje operona gena zaduženih za kasnije faze. Proizvodi ove druge serije gena moći će djelovati i u smislu izgradnje novih embrionalnih struktura i u smislu potiskivanja prethodnih operona i uklanjanja potiskivanja sljedećih, u uređenom slijedu koji dovodi do izgradnje novog organizma , zahvaljujući genetičkim informacijama prikupljenim iz genoma kroz tisućljeća u sve razvijenije vrste.
Haeckelov poznati izraz "ontogeneza sažima filogeniju" zapravo izražava činjenicu da više vrste ponavljaju, u fazama embrionalnog razvoja, sukcesiju koja je već pronađena u evolucijski prethodnim vrstama.
Rane faze embrija imaju tendenciju da budu slične kod kralježnjaka, osobito do pojave škrga.
Kod vrsta koje prelaze na zračno disanje, škrge se zatim ponovno apsorbiraju i ponovno koriste (na primjer za stvaranje endokrinih žlijezda), ali su genetski podaci koji se odnose na stvaranje škrga sačuvani i kod ljudi. Ovo je očito primjer embrionalnih strukturnih gena koji su prisutni u genomu svih kralježnjaka i moraju ostati potisnuti nakon što su funkcionirali u svom ontogenetskom trenutku.
Tumačenje embriogeneze u smislu regulacije djelovanja gena omogućuje objedinjavanje složenih tradicionalnih iskustava eksperimentalne embriologije.
BLIZANCI
Zigota i prvi blastomeri, sve dok ne počne sinteza proteina, totipotentni su, odnosno sposobni su oživjeti cijeli organizam. S tim su povezani eksperimenti Spemanna, koji je iz davljenja žigote vodozemaca dobio dva embrija. Sličan fenomen pojavljuje se u osnovi fenomena jednojajčanih blizanaca u čovjeka, koji se upravo iz tog razloga nazivaju monozigotnim (MZ).Spemannovi eksperimentalni blizanci bili su upola manji od normalnih, dok su kod čovjeka potpuno normalni.To objašnjava zašto su u vodozemaca dva embrija morala dijeliti jedini već dobiveni žumanjak, dok u čovjeka embriji mogu primiti, kroz posteljicu, sve što je potrebne za njihov razvoj i rast.
Treba se sjetiti da u "čovjeka dvije trećine slučajeva blizanaca" ima drugo podrijetlo: oni proizlaze iz povremenog istodobnog sazrijevanja dva folikula, s oslobađanjem dva ovala koji, kad se oplode, daju dva zigota; zapravo, u u ovom slučaju govorimo o dvojajčanim blizancima (DZ).
Budući da blizanci MZ, podijeljeni mitozom od pojedinačne zigote, imaju isti genom, razlike među njima moraju biti ekološkog podrijetla. Umjesto toga, genom dva blizanca DZ sličan je samo onoliko koliko i kod bilo koje dvije braće. Metoda blizanaca, naširoko koristi u humanoj genetici, ali i u području sporta.
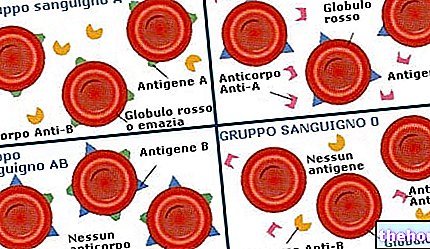
U "čovjeku, u kojemu bi određeni etički razlozi zabranjivali eksperimentiranje, može se ustanoviti koliko je neki karakter reguliran nasljednim čimbenicima: zapravo, strogo naslijeđeni karakteri (poput krvnih grupa) uvijek su podudarni samo u blizancima MZ; podudarnost svojstva u MZ bliska je onoj u DZ, zaključuje se da čimbenici okoliša prevladavaju nad nasljednim u određivanju te fenotipske osobine.











.jpg)